搜索

 首頁
首頁

 EON4概況
EON4概況

 師資隊伍
師資隊伍

 人才培養
人才培養

 招生就業
招生就業

 科學研究
科學研究

 平臺基地
平臺基地

 黨群工作
黨群工作

 校友之家
校友之家

 安全工作
安全工作

 網上辦事
網上辦事



 當前位置:
當前位置: 上海交大張大兵教授研究團隊經過5年的研究,克隆並鑒定了一個參與調控單子葉植物水稻花藥發育的堿性螺旋-環-螺旋轉錄因子(basic Helix-Loop-Helix transcription factore)💇🏼♀️,EAT1(Eternal Tapetum 1),並提出植物發育性的程序性細胞死亡新機製(如下圖所示)。該研究成果已經被《自然通訊》(Nature Communications)2013年2月5日發表😱。
程序性細胞死亡(Programmed Cell Death, PCD)是廣泛存在的一種死亡方式,在細胞分化和個體發育中起到至關重要的作用。植物基因組中缺乏動物PCD過程的效應蛋白——胱冬酶(Caspase),但許多植物細胞死亡過程表現為類似於動物PCD的行為🧘🏻♂️,例如染色質片斷化、線粒體異常👰🏽、細胞色素c(Cyt c)的釋放、caspase特異性底物活性反應等🧎♀️➡️。但是植物中程序性細胞死亡控製機理? 什麽類型蛋白可以發揮類似胱冬酶在PCD中的功能?
絨氈層是植物花藥最內層壁,在小孢子發育過程中,逐漸死亡👵🏼🪽,為小孢子發育提供營養和能量。絨氈層是植物發育性細胞死亡的理想的模式組織。通過遺傳學張大兵教授團隊研究發現了一個在絨氈層中特異性表達的bHLH轉錄因子🧖🏻♀️,EAT1,可以在轉錄水平上直接調節兩個天冬氨酸蛋白酶(Aspartic Preotease,ASP)的表達👩🦲。ASPs在酵母🪳、擬南芥和煙草體內過表達,會造成細胞和組織的死亡,說明ASPs具有廣譜的死亡促進者作用,而EAT1則是通過控製ASP表達執行死亡起始的作用♒️。體內和體外實驗證明EAT1蛋白能夠與調控絨氈層細胞死亡的另一個bHLH轉錄因子🙇🏽,TDR(TAPETUM DEGENERATION RETARDATION,Plant Cell,全文下載:http://www.ncbi.nlm.nih.gov/pmc/articles/PMC1693939/)直接作用。張大兵教授團隊2006年的結果證明,TDR通過直接調節一個半胱氨酸蛋白酶OsCP1(Cysteine Protease 1)的表達控製絨氈層細胞死亡。 基因表達分析結果表明TDR對絨氈層的發育調控可能部分通過EAT1實現; 有趣的是🦵🏼, EAT1不能直接調節OsCP1的表達🛀🏿👨🏻🏭,TDR也不能直接調節ASPs的表達。說明在絨氈層細胞死亡調控過程中🗳📤,EAT1和TDR可能存在著不同的分工😜。這項研究為全面認識花粉發育以及細胞程序性死亡等生物學過程提供了新的認識(http://www.nature.com/ncomms/journal/v4/n2/abs/ncomms2396.html?WT.ec_id=NCOMMS-20130205) 。
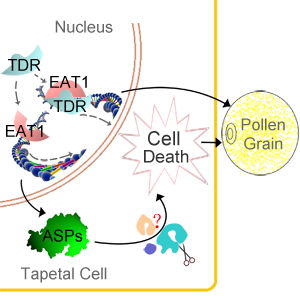
EAT1花藥絨粘層細胞程序性死亡作用機製的示意圖

EON体育4平台 Copyright © 2020 滬交ICP備51732244. All Rights Reserved.